基因敲除细胞系(KO)
立即咨询艾森生物依托成熟的AI-CRISPR™细胞基因编辑平台和完善的细胞培养体系,启动近5万个全基因组敲除细胞库构建计划,致力于为基础研究、抗体验证、药物筛选、疾病诊断、治疗和检测等方面提供大力支持。KO细胞库涵盖肿瘤、心血管、神经等20多种研究领域,现在下单现货KO细胞库,交付单克隆纯合子,快至1周发货,低至6880元,点击图片即可了解活动详情。
一、KO细胞敲除定制服务
作为实现基因功能缺失的重要调控方式,基因敲除(Knock out, KO)可完全消除目的基因的表达,使其蛋白完全不表达或功能彻底丧失,且可以更好地观察细胞表型的变化。但KO细胞株构建过程较为繁琐,实验周期长,且容易出现敲除效果不彻底的情况,导致研究进度受阻。
基于AI-CRISPR™细胞基因编辑系统,现在定制KO细胞服务,交付单克隆纯合子,快至30个工作日,低至10980元,满足您的早期筛选实验需求 。我们使用优化的转染体系将RNP(gRNA和Cas蛋白复合物)直接递送到细胞内,相比于质粒和病毒等CRISPR/Cas介导的方法,脱靶效率降低明显,能更精准地切割目标DNA序列。
基因敲除细胞服务详情,超300种细胞成功案例!
| 细胞类型 | 常规细胞系、肿瘤细胞系、IPS/ES等各类细胞 |
|---|---|
| 服务类型 | 单基因/多基因敲除 |
| 交付成果 | 纯合子克隆 |
| 周期/价格 | 快至6周,低至10980元;添加官方微信号:18520176000(手机同号) |
二、基因敲除细胞方案
艾森生物拥有多种敲除策略满足客户需求:我们具备移码突变、片段敲除、多基因敲除等编辑技术能力,根据研究项目的基因和细胞信息的不同,采取最佳方案,极大提高目的基因敲除的成功率和蛋白的不表达效率。
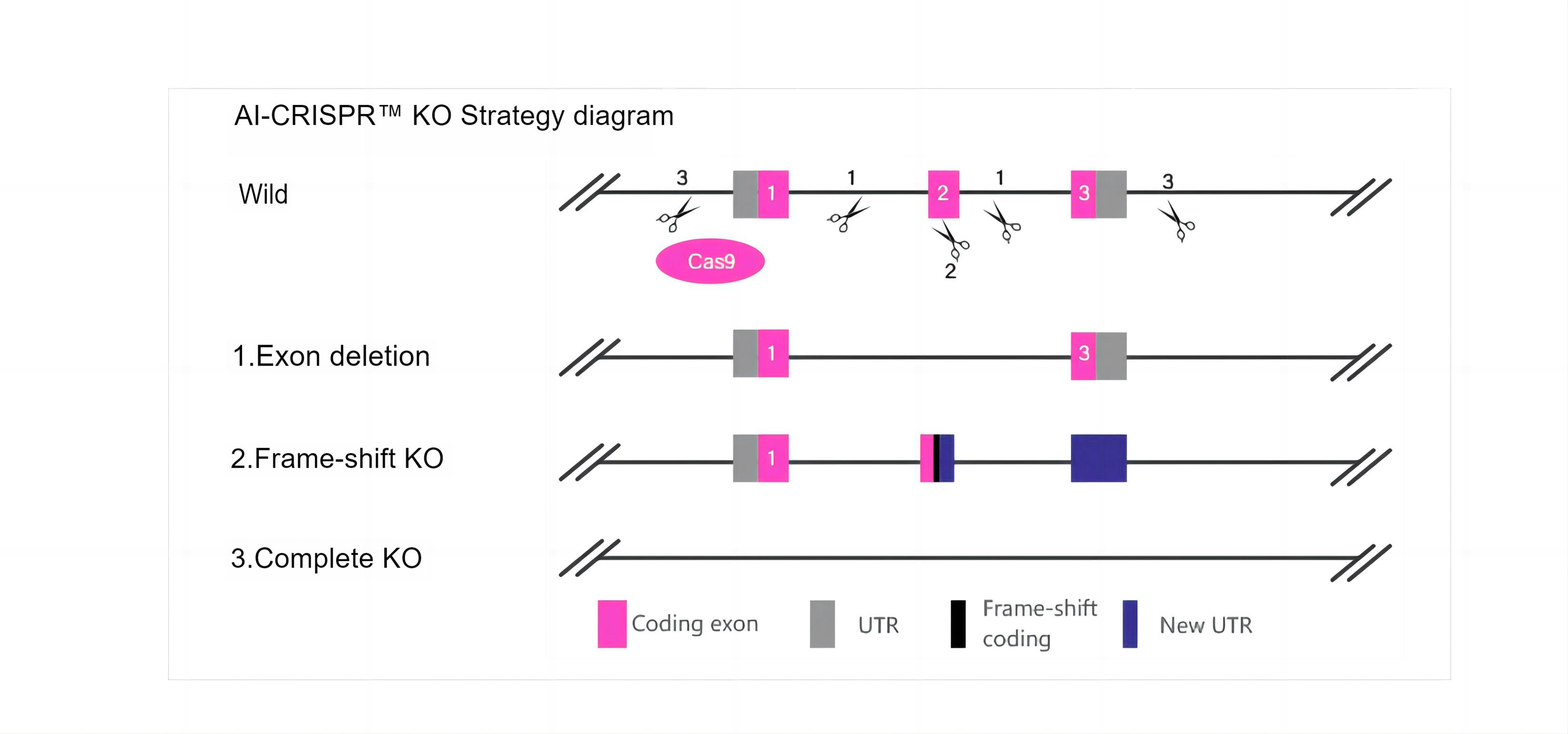
三、服务流程及质量控制
优化实验流程,有效提升编辑效率
基因/细胞分析→sgRNA设计与合成→细胞电转及KO效果鉴定→单克隆化→测序分析→复检及扩增。
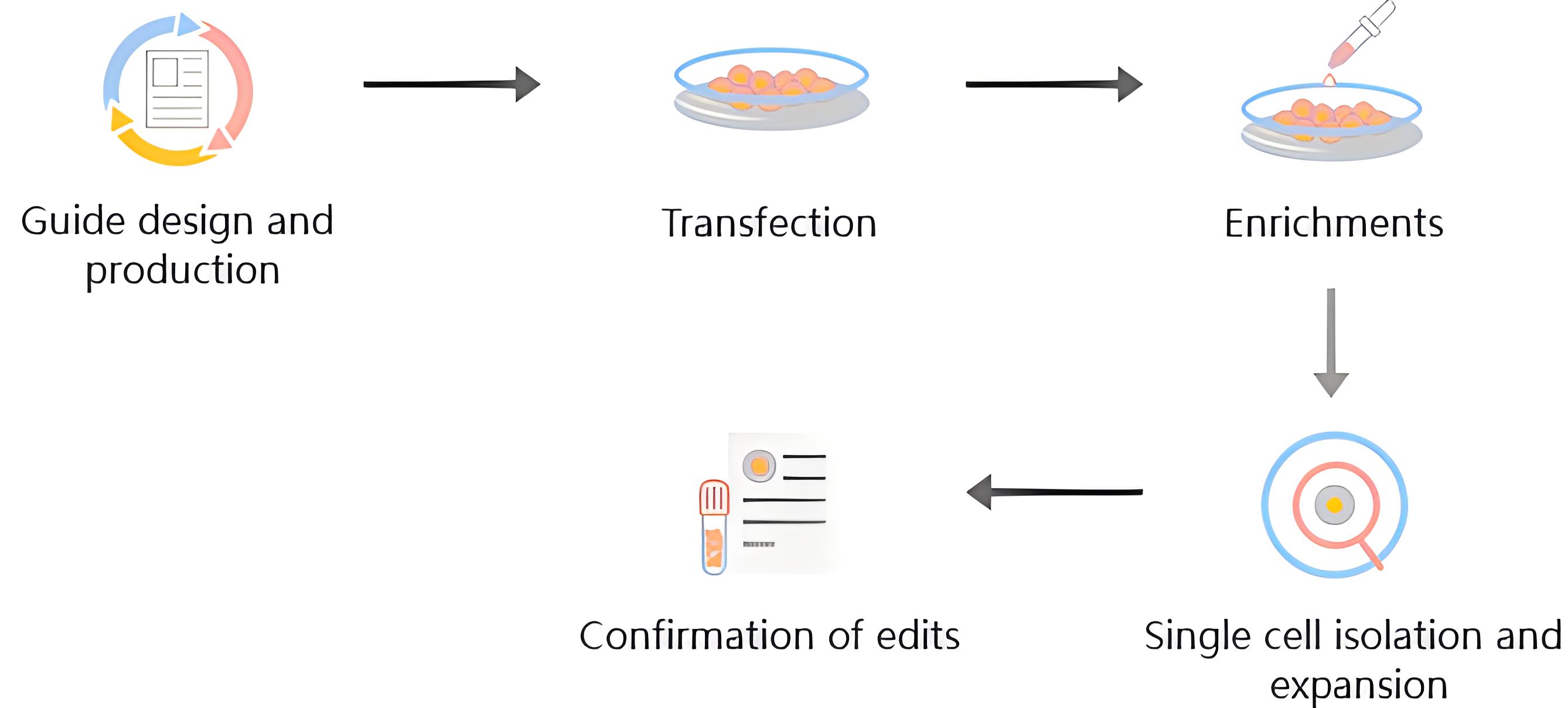
强大的算法赋能细胞的基因编辑项目——AI辅助筛选Western Blot阴性单克隆。
借助罕见病数据中心(RDDC)开发的RNA剪接模型工具,预测碱基序列突变后mRNA序列可能发生的剪切情况,辅助筛选ORF移码的单克隆,更好地规避在Western Blot环节检测出蛋白残留的风险。

四、AI-CRISPR™技术优势
1. AI-CRISPR™技术优势:拥有更高的基因切割效率与同源重组效率,基因编辑效率提升10-20倍。
2. 成熟的技术平台:大量成功案例,在超300种细胞中成功实现超数千例基因编辑;可提供从细胞基因表达调控、细胞功能验证及表型分析的全流程服务。
3. 脱靶效率低:采用RNP递送,相比于质粒或者病毒等基因敲除方法,具有更高的特异性和编辑效率。其能准确靶向目的细胞,提高效率的同时减少非特异的切割;实现Cell pool/纯合子交付,快至6周。
4. 应用广泛:AI-CRISPR™基因编辑轻松实现基因 敲除/点突变/敲入;承诺项目失败全额退款,成功保障安全无忧。
5. 严格的质量控制体系:进行细菌、支原体等微生物的双重检测,确保100%无污染,并充分鉴定基因编辑效果;进行细胞活率检测,保证交付质量。
五、案例分析
| 种属 | 人 |
|---|---|
| 细胞名称 | BEAS-2B细胞 |
| 培养体系 | DMEM-H+10% FBS+1% P/S |
| 基因型检测 | 针对gRNA打靶位置及其附近基因组序列进行测序。测序结果与理论序列一致 |
| 细胞转导 | 1350V,20ms,1次;电转体系:10 uL |
| 克隆形成率 | 细胞增殖能力较好 |
| PCR筛选 | 筛选克隆数量:170; 杂合子数量:13; 纯合子数量:2 |
| PCR方案1 |
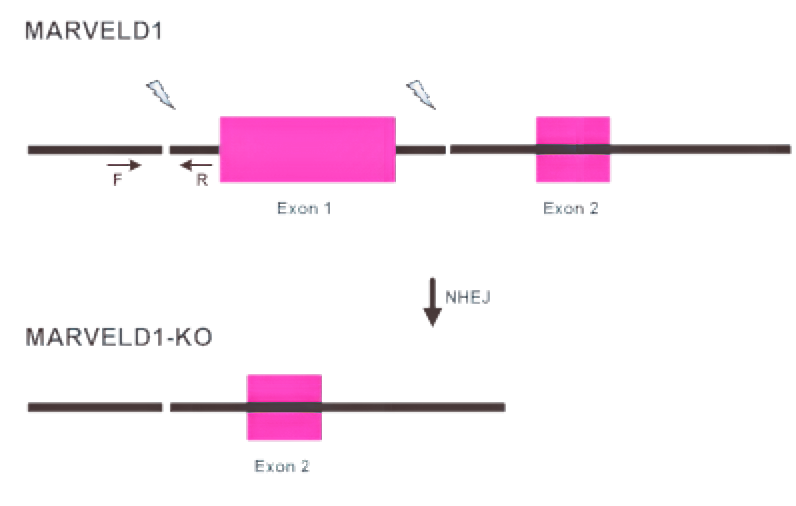
|
| 电泳图数据1 | WT:490 bp; 杂合子:490 bp; 纯合子:0 bp |
| 电泳图示例1 |
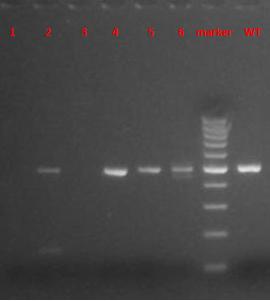
|
| PCR方案2 |
|
| 电泳图数据2 | WT:0 bp; 杂合子:~218 bp; 纯合子:~218 bp |
| 电泳图示例2 |

|
| 测序结果 | TAAGTCCGTGGGCTTTGCTTCCTCTTGGCCCTGGGTTTGTGTCCCAG-del-3310bp-TTTGCATGCTGGTCTCGAACTCCTGGGCTCAAGAGATCCTCC |
| 克隆状态 |
|
文献解读:CRISPR基因编辑BEAS-2B细胞系——希望就在呼吸之间
1. BEAS-2B细胞背景
BEAS-2B细胞是从肺癌个体的正常人支气管上皮病理切片分离出的上皮细胞。这个细胞引种自ATCC CRL-9609,又可被称为 支气管上皮细胞。BEAS-2B细胞系最初是通过使用腺病毒12-SV40杂交病毒感染,并通过连续细胞传代建立的永生化细胞系,它保留了对血清反应进行鳞关分化的能力,这种能力可用于筛选诱导或影响分化及致癌的化学或生物制剂,因此 BEAS-2B细胞被认为是研究药物代谢活化作用和呼吸系统肿瘤以及分子机制理想的细胞模型。
2. BEAS-2B的应用
BEAS-2B 细胞在成骨和成脂分化方面与 hMSCs 具有相似的潜力
将细胞分化诱导21天后,用油红染色分化脂肪细胞中的脂质液泡,然后使用茜素红染色分化骨细胞中的钙沉积。结果表明BEAS-2B和在成骨发生或脂肪发生方面均显示出几乎相同的阳性染色,这说明BEAS-2B 细胞能像hMSC1细胞一样能在诱导后表现出很强的骨细胞和脂肪细胞分化能力。
BEAS-2B可用于筛选化学和生物制剂诱导。目前,BEAS-2B已被广泛用于研究肺癌发生的细胞和分子机制,其中包括上皮间质转化(EMT)在肺癌和肺炎球菌感染中的作用等。此外,BEAS-2B细胞系已被用作体外细胞模型,用于检测和筛选具有潜在肺毒性或肺部致癌性的各种化学和生物制剂。
3. 使用AI-CRISPR™在BEAS-2B细胞中实现基因编辑
基因编辑和细胞模型的建立对于推进功能基因组学、信号通路、新陈代谢、细胞死亡、药物发现、药物反应和癌症研究等领域具有重要的意义。BEAS-2B细胞作为研究药物代谢活化作用和呼吸系统肿瘤以及分子机制的理想细胞模型,已经有许多研究人员利用CRISPR/Cas9技术对其进行编辑以研究癌症特征、耐药机制的披露、癌症治疗、细胞死亡研究、功能基因组学、信号通路、药物发现、药物反应和细胞治疗等多种具有重要意义的课题。艾森生物研发的AI-CRISPR™(基于CRISPR/Cas9技术)在DNA双链的切割上比一般的CRISPR/Cas9系统更有效率,不仅可以显著提高同源重组的效率,还能同时在体内外实现高效的基因编辑,对于BEAS-2B细胞的基因编辑实验有着极大的优势。
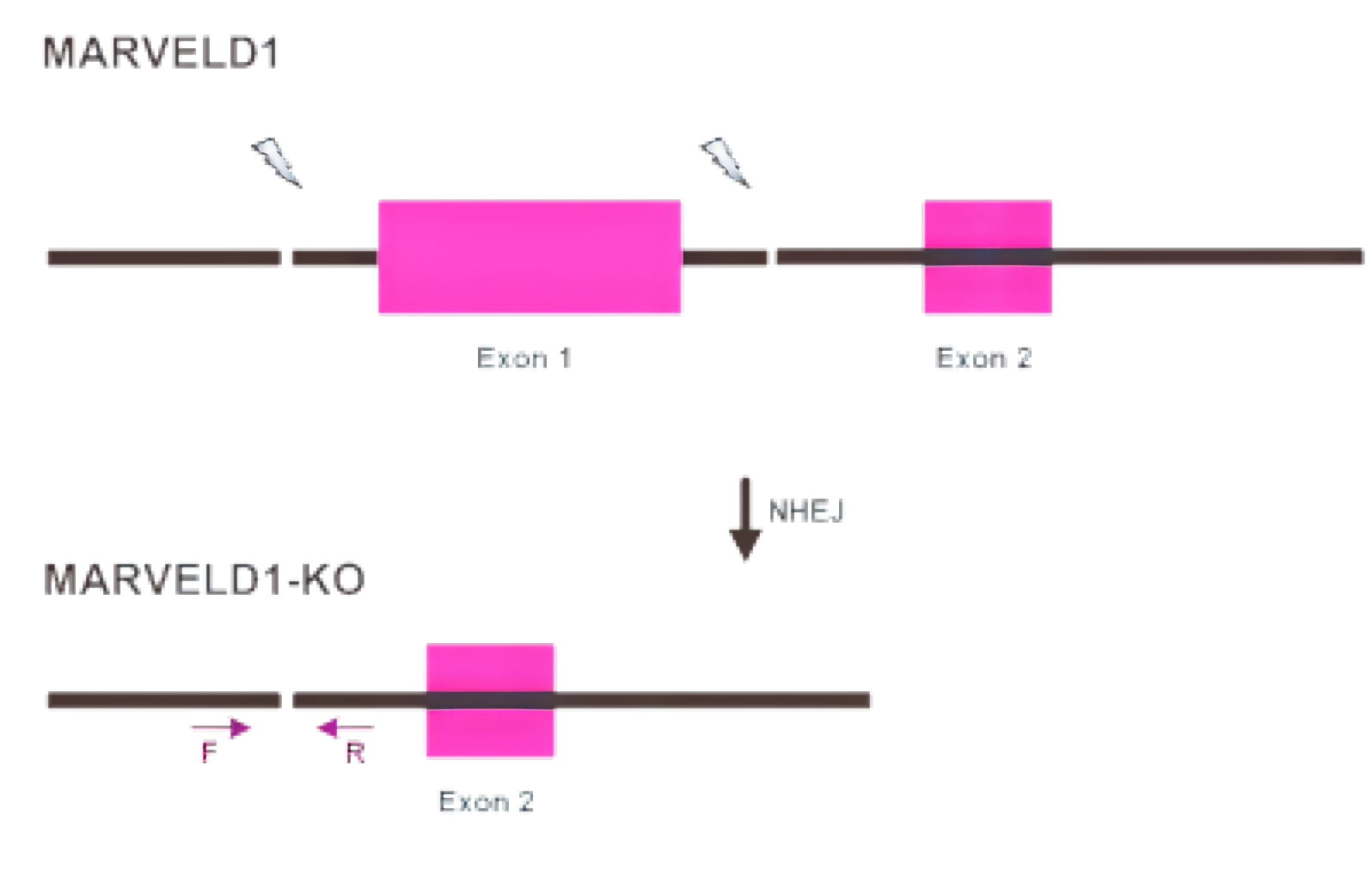
图1 使用AI-CRISPR™在BEAS-2B细胞中基因编辑过程
3.1 AI-CRISPR™基因敲除BEAS-2B细胞系
通过病毒转导、脂质体转染或核转移将gRNA和Cas9转移到细胞中,然后通过药物筛查后,产生单克隆,最终阳性克隆将通过测序进行验证。具体案例如下:
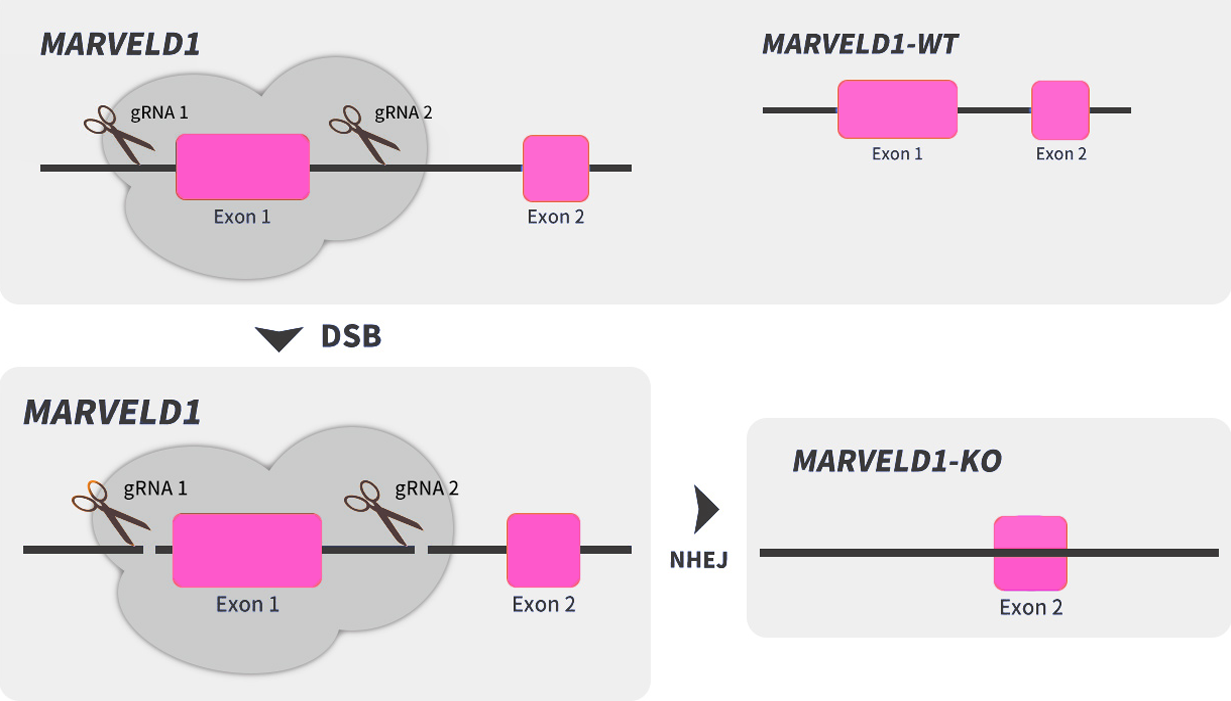
图2 BEAS-2B细胞系的敲除策略
P53基因是氡照射致BEAS-2B细胞恶性转化中的重要影响因素。在人正常肺上皮细胞(BEAS-2B)中 敲除P53基因,并对基因敲除后的BEAS-2B细胞和正常BEAS-2B细胞分别进行氡染毒处理。研究人员发现,BEAS-2B细胞敲除P53基因后氡照射的BEAS-2B细胞更早地出现了恶性转化。相较于BEAS-2B细胞染氡组(2B-Rn),P53基因敲除的BEAS-2B细胞染氡组(P53 KO-Rn)增殖速度更快(P<0.05),形成克隆的时间更早(P<0.05), 侵袭力更强(P<0.05),上皮-间质转化(EMT)相关基因mRNA表达水平变化也更早(P<0.05)。结果表明,氡照射导致BEAS-2B细胞发生恶性转化,并且P53在氡气致肺癌中具有重要作用。

参考:http://www.paper.edu.cn/releasepaper/content/201910-45.
3.2 基因敲入或基因点突变BEAS-2B细胞系
BEAS-2B细胞通过电穿孔法与gRNA、Cas9和Donor载体共转染,然后挑单克隆。阳性克隆将通过测序进行验证。具体案例如下:
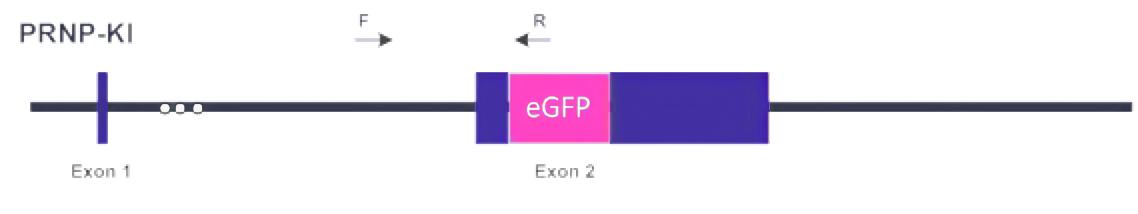
图3 BEAS-2B细胞系的敲入策略
(1)敲入案例
为了研究突变体NRASQ61K在血管再生中的特异性影响,在正常(非肿瘤)人支气管上皮细胞(BEAS-2B)中进行了敲入。与野生型(WT)细胞相比,在CAM上生长的NRASQ61K 过表达细胞的特征是血管系统紊乱,并存在若干个出血区域。为了证实体内NRASQ61K的促血管生成作用,在裸鼠身上进行了Matrigel plug试验,结果如图5B中所示,NRASQ61K表达细胞显示出血管再生明显增加,呈亮红色。然而, 野生型细胞中的血管再生可以忽略不计,并且血红蛋白水平较低。除此之外,研究者还发现NRASQ61K在体外的表达显著增加了类毛细管网络,如图5C所示。而肿瘤血管生成的关键所在正是细胞在骨髓上自发形成毛细血管样结构,因此,NRASQ61K的表达会在一定程度上促进肿瘤血管的生成。
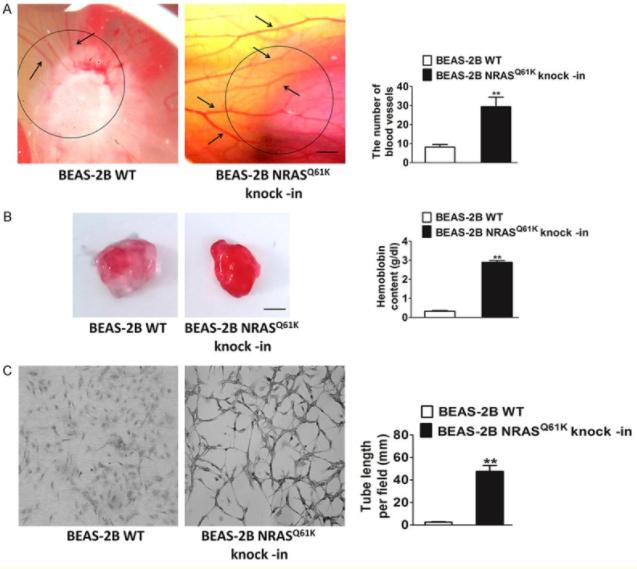
图5 将NRASQ61K敲入BEAS-2B细胞
(2)点突变案例
BEAS-2B细胞通过电穿孔法与gRNA、Cas9和Oligo共转染,然后挑单克隆。阳性克隆将通过测序进行验证。具体案例如下:

图4 BEAS-2B细胞系的点突变策略
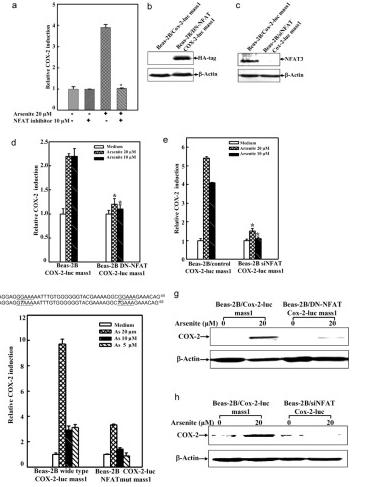
COX-2基因启动子区域包含NFκB、AP-1和NFAT结合位点,它们可以被这些转录因子识别,进而导致COX-2的转录。为了确定NFAT是否通过直接结合COX-2启动子区域来调节COX-2的表达,研究人员应用CRISPR/Cas9技术,对COX-2-Luc启动子区域的两个NFAT结合位点进行了点突变。结果表明,这种突变导致亚砷酸盐暴露诱导的COX-2转录受损。并且研究人员发现,与COX-2诱导相比,亚砷酸盐在研究中的NFAT激活达到了更早的峰值。因为亚砷酸盐的暴露只会导致BEAS-2B细胞中的NFAT的激活,而不是AP-1和NFκB的激活,所以亚砷酸盐激活NFAT是BEAS-2B细胞中COX-2诱导的原因。
另外为了证实NFAT3 在 BEAS-2B 细胞中亚砷酸盐诱导的 COX-2 表达中的重要作用,研究人员构建并使用了siNFAT3,他们将细胞用10μm NFAT 选择性抑制剂预处理,然后暴露于20μm亚砷酸盐中,研究人员发现BEAS-2B细胞中siNFAT3的稳定转染导致BEAS-2B细胞中NFAT3蛋白表达的明显降低(图. 6 中b)。siNFAT3对NFAT3表达的特异性抑制阻断了亚砷酸盐诱导的COX-2转录和蛋白质表达(图. 6 中e和h)。
这些结果表明了,NFAT3是BEAS-2B细胞中亚砷酸盐诱导COX-2的主要介质。
参考文献:
[1]Liu, Shuang, et al. "miR-106b-5p targeting SIX1 inhibits TGF-β1-induced pulmonary fibrosis and epithelial-mesenchymal transition in asthma through regulation of E2F1." International Journal of Molecular Medicine 47.3 (2021): 1-1.
[2]Sawada J, Li F, Komatsu M. R-Ras protein inhibits autophosphorylation of vascular endothelial growth factor receptor 2 in endothelial cells and suppresses receptor activation in tumor vasculature. J Biol Chem. 2015;290:8133–8145
[3]He, Jun, et al. "Arsenic-induced metabolic shift triggered by the loss of miR-199a-5p through Sp1-dependent DNA methylation." Toxicology and applied pharmacology 378 (2019): 114606.
[4]Pircher A, Hilbe W, Heidegger I, Drevs J, Tichelli A, Medinger M. Biomarkers in tumor angiogenesis and anti-angiogenic therapy. Int J Mol Sci. 2011;12:7077–7099.
[5]Song, Zhaowei, Fenghai Liu, and Jie Zhang. "Targeting NRASQ61K mutant delays tumor growth and angiogenesis in non-small cell lung cancer." American journal of cancer research 7.4 (2017): 831.
[6]W.R. Chappell, B.D. Beck, K.G. Brown, R. Chaney, R. Cothern, C.R. Cothern, K.J. Irgolic, D.W. North, I. Thornton, T.A. Tsongas. Environ. Health Perspect., 105 (1997), pp. 1060-1067
[7]Ding, Jin, et al. "Cyclooxygenase-2 induction by arsenite is through a nuclear factor of activated T-cell-dependent pathway and plays an antiapoptotic role in Beas-2B cells." Journal of Biological Chemistry 281.34 (2006): 24405-24413.
[8]H.Y. Chiou, Y.M. Hsueh, K.F. Liaw, S.F. Horng, M.H. Chiang, Y.S. Pu, J.S. Lin, C.H. Huang, C.J. Chen. Cancer Res., 55 (1995), pp. 1296-1300
[9]Van Den Heuvel, Rosette, et al. "Identification of PM10 characteristics involved in cellular responses in human bronchial epithelial cells (Beas-2B)." Environmental research 149 (2016): 48-56.